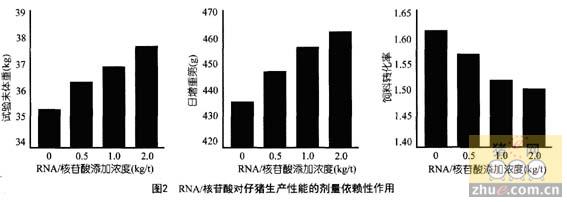
�����������о���չ
����: 2010-04-03 | ����: ���� | ��Դ: �Ѽ�
�߲�Ŀ��ͨ���������ֿ��Խ��͡�ͬʱ�����������ر��Dz����Դ�Ⱦ����������в���Ľ���������Ԥ�����ַ�������Ҫ�ķ������ã���δ����ȫ���ƺ�����Ⱦ�������С��ӳ�Զ�����������Ŵ�ѧ�������Ŵ�������������Բ�ԭ�Ŀ��ԣ���չ�������־����α��Ĺ�Ч�� 20����30���������ѧ�߱����˲�ͬ��Ʒ�ֶ��������ϲ������Դ��ڲ��죬��������һ��ʱ���ڶ������������ݿ������ֽ����˽�Ϊϵͳ���о�����������ѧ��������Ʒѧ�ķ�չ���ִ�Ⱦ����ͨ��Ԥ�����ֵõ�����Ч�Ŀ��ƣ�ʹ���������ܵ�һ�������䡣80������������ŷ�������ѧ�ͻ��̼����ķ�չ��Ϊ���������ṩ���µ�˼·��Ҳʹ�����������ӶԿ����������о���
����1 ���������Ŵ����� ��ԭ���ڴ�Ⱦ�����У�������3���������ƣ�����Ƥ�������ơ��������Է������ƺ������Է������ơ��������ܵ���ԭ����Ⱦʱ���������3����ķ������Ƽ��Եֿ������Ƿ�ȡ������Ⱦ�ͷ�����������õĽ�����������ܼ�ǿ��������ֳ���Ȼ�����������������Ŵ������IJ�ͬ�ɷ�Ϊ���⿹������һ�㿹���������⿹������ָ����ij���ض�������ԭ��Ŀ��ԣ����ֿ�����Ҫ��һ��������λ����ƣ�Ҳ�ɳ̶Ȳ�ͬ��������λ�㣨���������ӣ�����������Ӱ�졣�����е��о�������������⿹���������ڻ��������������ڴ��ڻ�ȱ��ij���ӷ��ӻ�����壬���ַ������������ã��پ�������ʶ�����������巴Ӧ�ڣ��ھ�����ԭ������⸽�������۲�ԭ��������ں���������ֳ�������ܷ�������������һ�㿹���������ڿ�ijһ�ֲ�ԭ�壬���ܶ����ͻ������ۺ�Ӱ�죬��ԭ��Ŀ�ԭ�Բ����һ�㿹����Ӱ�켫С����������û��Ӱ�죬���ֿ����������˻���Լ���������������ܡ��缦��mhc�������˲�����没����˹�����Ȳ��Ŀ��Ժ��������йء�
����1.1 mhc�뿹������Ҫ��֯�����Ը����壨major hlstocompatiblllty complex�� mhc�����뿹���Ժ�����Ӧ��������ص�һ�����Ⱥ������ϸ�����������Ե��ף����������нϸߵȵĶ����С�mhc����3�������Щ�����ڳ�Ⱦɫ���Ͻ��������γɵ����ͣ��ʳ�Ⱦɫ�干�����Ŵ��� ����mhc������Ϊsla��λ������7��Ⱦɫ���ϣ�warner��1987��������1�ͺ͢��ͻ�������1�ͻ�����м�ǿ�Ķ�̬�ԡ�sla������������Ӧ��������أ�roths��hild�ȣ�1984��֤ʵ������5�����ֶ�֧���ܰ�Ѫ���ϸ˾�������Ӧ����sla�Ŀ����¡�mallard�ȣ�1989����nih �����з��֣�sla������dd��dg��ss����������ϸ����ĸ�������ܾ�ø�ͺϳ��ģ�t��g����a��l��ǿӦ���ߡ��ڼ������Է��棬tissot���ȣ�1989�������Ŵ���Ƥ�����Ժ�����sla�������йء�renard�ȣ�1985�����ָ�к��ɵĶ���ǰ��������sla�ĵ����������йء�����sla�IJ�ͬ�����ͶԼ�����Ŀ���Ҳ�������Ͳ��죨lunney�ȣ�1988����warner��1991���Ƚ��������о����뼲������ϵ�Լ�sla������Ľ�Ϻ��ܣ�lunney�ȣ�1998����valman�ȣ�1998���ֶԴ˽����˸��¡�
����1.2 ���������� ���⿹�����ɶ�ij�ּ�����ԭ������ض����Ե���������ƣ�ʶ������ض��Ŀ�������Կ������־����ش�����壬��������ֱ�ӽ���������ѡ�����֣����ҿɽ�һ������ת��������������о��ý�����������˾�k88��f18������� ����������к������k88�����ϸ�����濹ԭ������С��ճĤ��Ƥϸ��������Ӧ�����������𣬴˾�������Ӻϴ�����ֳ���ͷų����ض����¸�к��ȱ��k88����������ܿ�k88���ţ��Ӷ��ܿ���к��sellwood��1975����k88�������λ������13��Ⱦɫ���ϣ�rothschild,1992�����������������һ�Ե�λ����s��s���ƣ������壨s���������壨s����ȫ���ԣ����ϵ¶��Ŵ���k88��������Ƶ��������������ܸߣ���Ĵ�����k88������ռ���о��Ĵ�ͳ�����Ⱥ���88����sondgrasss��1981�������й��ط���k88����Ƶ�ʷdz��ͣ�������Ȼ�Ŀ����ԣ�chappuls��1984����bertschinser��1983�����ֶ���ǰ�������ˮ�ײ���f18������ϸ�������������ͷų����ض�����ģ�ȱ��f18��������������ܿ���к��ˮ�ײ���f18�������λ������6��Ⱦɫ�壬����Ѫ����������s����ϸ��øϵͳ����������������������Ϊ����f18ճ���ؽ�ϵ��������Ϊ�����Ի��������Ŵ�����֮��Ϊ���Ի��������Ŵ���vogeli�ȣ�1992����vogeli��1997�����ú�ѡ���������������о��˶�λ��6qll���ε�a��1��2�������ǻ�ת��ø����futi��futz����Ϊ����f18ճ����1����ѡ�����������뿹�����йص�������Ͷ�λ��13q31���ε�e��colt f4�������Ҳ�����ĸ�к�йء�
����1��3 ������������Ӧ����������״�Ĺ�ϵ ������������Ӧ����������������֮���ϵ��ʵ��֤�ݻ���ȱ�������Ҳ��һ��vander�ȣ�1983���Ը������ʵ�ѡ������˶��������ϲ������ԣ�lamont�ȣ�1989��ȴ�����������ϲ��Ͳ������ܼ�Ĺ�ϵ�ʸ���ء������У�meeker�ȣ�1987�������������������֧���ܰ�Ѫ���ϸ˾���Ʒ�����α��Ȯ��������������Ӧ��ʸ���أ����г���k88������������Ͽ죬���������ʽ��ţ�edfors�ȣ�1986��������sla�����������״����������warne�ȣ�1991�� lunney���� 1998������Щ���ͬ����sla����������Դ�ͷ�ֳ��״�����ϵ�������أ�����sla����Ӧ���ǽ�����ֵ���ѡ����mallard�ȣ�1998���ھ���8��ѡ���γɵ���������Ӧ��hϵ���͵�����Ӧ��lϵ��Ʒϵ�з���hϵ��lϵ��10d�ﵽ�������ء�
����2 �������� ���ڶ��ּ�����������Ŵ����죬�ӳ�Զ����������ѡ�������п�չ�����������Ŵ���������߶Բ�ԭ�Ŀ��ԣ���������߽������������ļ��������ڿ�չ�������ֵķ����У�������Ȼ�����������˹������¶�������״����ѡ��������ѡ����Ӧ�Ŀ�����״��ahlborn��breier�ȣ�1990�������÷���������������״�Ϳ�����״������ԣ�һЩ������״�Ϳ�����״������ϵ����С�������ʸ���ء��ڽ�������״��Ϊ����״���ֶ����һ���֣�������ϲ����ۺ�����ֵ����Ϊ����ѡ������ݣ�warren�ȣ�1990������ȱ�������Զ��κο�����״�ľ���Ȩ�ؽ��ж��壬ͬʱ������״��b�����ӣ�ѡ���չ������١�
����2��1��1 ���ݼ�¼����ֱ��ѡ�� �÷�����ֱ�ۡ�ȷ���ŵ㣬����Ⱥ������û��Ӱ�죬ȱ������Ⱥ��������ѡ����ѡ���Ч��ֵ�û��ɡ����淽����ֱ�Ӷ����ָ��塢ͬ���ͺ�����й���ѡ�����ַ�����ȱ�������ɷ���̫�ߣ�ȡ���ڼ��������ij̶ȣ���ͬʱ��ֱ��Ӱ����Ⱥ���������ڸ����ַ������缸��������һ���̶ȵļ��������DZ���ģ���Ϊ����ʱ��ѡ��������ұ��������ģ��������ܲⶨȷ�Խϲbarker�ȣ�1990��blup���������ܲⶨ�����ͬ���ͺ���ⶨ�����ѣ��о�������Ժ�ѡ�����¡���й������飬���ַ���ȷ�Խϸߣ�������û��Ӱ�죬����ѡ��Ч�����ԡ������������¡�����ɴ��ۺܸߡ�
����2��1��2 ���Կ˷���ȱ�� �ټ�����״���Ŵ����ϵͣ��Ҳ�����״������״��ֱ��ѡ���ȷ�Բ�ں��̲ⶨ�Ѷȴ����������������Щ������״���Ӻ����ԣ��е������������ض����������Խ���ȷѡ������ȷ����ͬ�����������ij̶Ⱥͱ�������Щ������״��������״���ڸ���أ����Խ���ѡ��
����2��1��3 �������ļ��ѡ�� ����ֱ��ѡ����ڵ����ѣ�����ѧ�߳��ԶԿ������ֽ��м��ѡ�������ļ��ѡ������2������Ӧ�������������Ϊһ�ֿ���ѡ��ļ��ָ�꣬���ڵ��о�������С��biozzi�ȣ�1980���ͼ����У�lamout�ȣ�1990���������߷���buschmann�ȣ�1974��1980���ֱ��������ϸ���Ͱ뿹ԭ��dnp��hapten����Ϊ�������ʣ���ʾ�����ּ��Ʒϵ�����Ҫ���졣rothschild�ȣ�1984����֧���ܰ�Ѫ���ϸ˾���Ʒ�����α��Ȯ����������н��֣��������ּ��Ʒϵ�����������������Ӧ��ʵ͵��е��Ŵ�����0��05��d��52����edfors�ȣ�1985����2��ecoli��ԭ������Ӧ��Ҳ�����е��Ŵ������Ŵ���Ϊ0��29��0��45�� </p><p>����2��2 �Ŵ���Ǹ����������� ��λ�뿹����״����������dna���ӱ�ǣ�ͨ����ӶԱ��ѡ���ﵽ����ѡ���Ŀ�ģ����ַ�����Ҫ��ȷ��������״���Ŵ���ǡ�
����2.2��1 ��Ǹ���ѡ��mas�����������Ǽ����������ϲ���mhc�Ĺ�ϵ��b19���ԣ���b20�У��ɼ��ͨ��ѡ��b21���ϸ���������ò�����Ⱥ������ʵʩ��ȷ�Խϸߣ��ɱ��ͣ��ɽ�������ѡ���ǵ�ǰ�����н�ʵ�õķ��������ñ�Ǹ���blup���ɽ�һ�����ȷ�ԣ��ӿ��Ŵ���չ��Ŀǰ��Ӧ������mas���Ŵ������Ҫ�Т�mhc�����ͣ�mhc��Ϊ���ӱ�ǽ��и���ѡ��ͬʱ�Զ��ּ�������ѡ��һ�㿹����ѡ��Ч���Ϻá����⣬mhc�����������״������������������߿���������������ͬʱʹ�������ܵõ���ߡ���futi��ǣ�vogeli�ȣ�1997���Ըû��������pcr��rflp�����������ϸ����ճ�����鷢�ָû����307λ��߶�̬�ԣ�����f18����λ����ڸ߶�������ƽ�⣬307λ�������g��a��ͻ�䣬�Ӷ�ʹa����ʧȥ�� 1��hin6��ø��λ�㣬����3�ֲ�ͬ������������aa������������Ϊ���ԣ�gg�����ͱ������С�meijerink��1997���Դ˱�ǣ���250ͷ��ʿ��Լ�ˡ�����ˡ������ġ�Ƥ�������������л����Ͳⶨ�����ݲⶨ���ͨ���ʵ���ѡ��ʵʩ�˿������֡�����ars���´˳ɹ������ģ��ҵ��������1999��7�������ڴ�Լ�������ɸò��Ŀ�������
����2.2.2 ��Ǹ������루mai�� ����;���У��������Ŵ���ǽ���Ŀ�Ļ������룻�������Ŵ����ѡ�������͡�����ʱ��hanset�ȣ�1995��ͨ�������ryri�������������������칹ø��gpi��λ�㣬��������������Ի������뵽Ƥ�����У�����3�λؽ�����÷�������Ƥ����Ʒϵ������mai�����й��ط�����������ǿ����ȷ��������������ͬʱ�������������������ԡ�
����2��2.3 ֱ�ӽ���������ѡ�� ���ijһ�ض�������Ū�忹�������Ŵ����ƣ�ֱ�ӶԿ�����������л�����ѡ���Ч�������롣�Ժ���̽��Ϊ���������÷����ӽ���ӫ������Ӱ�ķ������������Կ��ٰ�ȫʶ���������������k88���Ի���8�ı�Ǻ�ѡ�Ϳ��Լ��������ͳ����ss�������������ǾͿ��Է�������k88���п��Եĺ����Ӣ���о��߷������Ժ����Ի���֮��IJ���ֻ����ͬһλ�õ�3�������ͬ���ݴ����Ƴ�dna̽���Լ���������ѪҺ�Ϳ�ȷ�ؼ��Ӧ������Գ��
����2��3 ����ת���̿������� �Ӷ�ʮ����80�����ʼ�����dz��Խ���ת���̿������֣����ݿ��Ի���ת�Ƶ�������IJ�λ���ɽ����Ϊ2�ࣻ�ٺ������ߣ�����Դ����ע�䵽������ϸ���ڣ�ʹ������ϸ���ڱ��ﲢ��ÿ��ԣ����ֿ��Բ���ͨ�������������������ת�������������Դ���������������Ŀ�¡Ƭ��ת�Ƶ��������ڵ������ϱ���Բ������µ��Ŵ���������״�Ķ�����ܽ��µ��Ŵ���Ϣ�ȶ����ݸ���������м�ֵ�ĺ�ѡ������������ػ�����������������в�����������ᡢmhc����ø�������¿ǵ�����Ͳ����к��Ե���¡���������Щ����Ŀ�¡Ƭ�ο�ͨ��ϸ����ע�䡢�������巨����̥��ϸ���齨����ϸ����¡����ת¼�������巨�Ȼ�������������ϸ���ڻ�ñ��ʹ���Ŀ���������ǿ�������ض��Ŀ�����Ⱥ�������������ɹ����в�����mx����ת���������ҹ��ѽ����Ό�����������롰863�ƻ��������ɹ����ת�ƿ�����������ø��ת�����á�
����2��4 ��չ�������ֵ����Ѻʹ������� �ٿ����Ե��Ŵ����Ʒdz��������ܻ���Ӱ��ϴڲ�������Ŵ����Լ���������������ϵҲʮ�ָ��ӡ��ۿ����Ի�����ָ�����Բⶨ����ȱ�����м��ѡ��Ŀɿ���ǡ��ܲ�ԭ�������Ѹ�٣����γ��ܿ˷��������ı���Ʒϵ���ݿ�������������״֮����ڸ���أ���ͬ������Ҳ��������ԡ�����������ϳ������뾭����ѡ��
����3 չ�� ���ںܶ����������˳������ַ����ڿ��������е�Ӧ�ã������Ŵ���Ǹ������ֺ�ת�������ֽ��ǽ�����ֵ���Ҫ��������sla��������Ҫ�ĺ�ѡ���� Ŀǰ�����������йص�������״��qtl������λ�о��ý��٣��ҿ��Ի���ļ���������Ϳ�¡�������д����ơ������յ��뿹����״�����������Ŵ����Ҳ���٣���ȱ���ɿ����Ŵ���ǡ�ת�������֣���������Դ�����ѡ��ͷ��롢����ĵ��뼼���ȷ��滹����д�����̽���Թ���������ת�������ڰ�ȫ�ԡ���̬ѧ������ѧ�����������������⡣�����ŷ�������ѧ�ͻ��̼����Ľ�һ����չ�Լ���������DZ�ڵľ�ЧӦ������ʹ�������ֳ�Ϊ21���������ֵ�һ����������