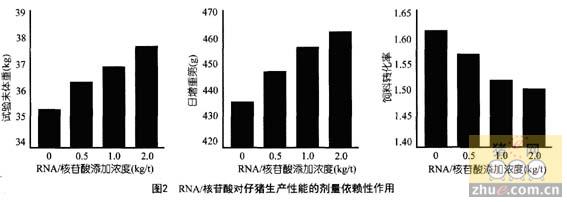
寡肽的转运机制及其影响因素
发布: 2013-09-29 | 作者: | 来源:
1寡肽的转运机制
完整肽进入上皮细胞,而在细胞内水解的吸收通路的存在被忽视了相当长的时间。早在100多年前就有人提到了肽转运的可能性(Matthews,1987[5])。Agar(1953[6])年证实了完整双甘肽在大鼠肠道跨上皮的转运。但是由于受传统蛋白质消化吸收理论的影响,学者们对其它的吸收方式不容易接受,并且由于双甘肽被认为是一种特殊的二肽,它的分子量很小,因此这一发现的重要性没有被认识到。直到20世纪60年代出Newey和Smyth(1959[7],1960[8])第一次提供了肽被完整吸收的资料。他们发现。蛋白质在小肠中的消化产物不仅有氨基酸,还有大量的寡肽,而且肽可完整的进入肠粘膜细胞,并在粘膜细胞中进一步水解生成氨基酸进入血液循环。以后,1965~1980年间,积累了越来越多关于完整短肽肠道转运的证据。尤其是伦敦的David M.Matthews和Pittsburgh的Siamak A,Adibi研究小组证实了肽的转运可能性不仅具有学术意义,并且可能代表着一个与相应的游离氨基酸吸收同等重要的氨基酸氮摄入的吸收通路(Matthews and Adibi,1976[9]Matthews,1987[5],1991[10])。肠粘膜对氨基酸和肽的吸收过程是复杂的。一般认为二肽、三肽被吸收摄入肠细胞后,被肽酶水解以游离氨基酸的形式进入血液循环。但近年的营养生理和药理试验证实,在某些情况下完整的肽能够通过肠粘膜的肽载体进入循环。乐国伟(1995[11])给来航鸡肠道灌注酪蛋白水解物后,用高压液相色谱检测寡肽的结果表明,肝门静脉血液和肠道内容物中出现了相同的肽类物质,由此推断,鸡肠道可吸收完整的肽,并能进入血液循环。动物体内可能存在多种寡肽的转运体系,寡肽在不同种动物体内的转运方式可能不同,同种动物也可能有集中不同的寡肽转运方式。
1 .1单胃动物寡肽的转运机制
日粮蛋白质在动物消化道内经过肽酶的作用,降解为游离氨基酸和寡肽,寡肽在小肠绒毛膜刷壮缘受到氨肽酶A和氨肽酶N的作用,最终分解为游离氨基酸和小肽的形式,被动物利用。目前,关于寡肽的转运机制还不完全清楚,还处在进一步的研究探讨中。Daniel等(1994[12])提出关于小肽转运进入肠上皮细胞的假设模型。其假设为:在正常生理条件下,肠上皮细胞处于一种外正内负的膜电位极化状态,并在细胞膜两侧保持着一种较强的质子电化学梯度。同时,细胞内各种肽分解酶使细胞内保持着较低的肽浓度。当消化道内饲料蛋白被消化酶分解释放出大量小肽时,在肠细胞膜内形成了短时间的小肽浓度梯度。在该浓度梯度以及由膜电位所形成的质子电化学梯度的驱动下,吸附在细胞上的H+和小肽借助共同的肽/H+偶联载体向细胞内进行跨膜转运。运输到细胞内的小肽被迅速分解成游离氨基酸供细胞利用或转运到血液或其他细胞。而经共转运进入细胞内的H+引起细胞内pH值的降低,从而激活细胞膜上的Na+-H+互转通道,泵出H+,摄入Na+,从而保持细胞内pH值的稳定。同时细胞膜上的Na+-K+泵开放,泵出3个Na+,摄入2个K+保持细胞膜内Na+、K+正常的比例。,维持细胞内高K低Na的微环境。在整个过程中,细胞内各种酶对肽的降解作用,细胞内游离氨基酸的利用和转运,以及Na+-H+互转通道和Na+-K+泵对离子的转运共同维持着细胞膜内外小肽浓度梯度和质子电化学梯度,从而保证小肽跨膜运输的持续有利的进行。由该模型假设表明,小肽转运可能是一种浓度和pH(质子电化学梯度)依赖性、肽/H+共转运的载体被动运输过程。有试验为该假设提供了一定的依据。Gamapathy等(1981[13])、Rajendran等(1985[14])研究了兔、大鼠、人的肠道刷壮缘膜囊(BBMV)发现即使缺乏任何浓度梯度,肽的摄入仍为肽浓度的函数,并遵循米氏动力学常数,这表明一易化扩散系统参与了肽的膜转运。在这种情况下,只有驱动力是向内的底物浓度梯度。只要转运进入肠细胞的肽被水解,这种易化扩散将不断的维持。H.Chen等(2000[15])对鸡肠肽载体的研究表明,转鸡肠肽载体基因的中国仓鼠卵母细胞对肽的转运速度随Gly-Ser浓度的升高而升高。Daniel(1988[16])用pH微电极证实了小肠细胞内外pH值梯度的存在,其试验结果表明,即使肠腔内pH值为7.4,肠绒毛上部吸收区域的pH值仍为6.5~6.7,细胞内pH值为7.2~7.4,BBMV内外pH值梯度则为0.5~0.8pH单位。Tnwaites等(1993[17])用载有对pH灵敏的荧光染料的CaCo-2细胞(来源于人结肠肿瘤的细胞,在多孔流性支持物上发育成具有小肠上皮细胞的典型特征)研究肽转运对细胞内pH值影响的结果表明,随CaCo-2细胞外Gly-Ser浓度的增加,该细胞内pH值呈下降的趋势。,而细胞内灌注无肽溶液时,细胞内pH值则有所上升。这些试验都在一定程度上为肽载体转运假设提供了依据。此外,另有试验表明,肽的转运还有以下两种可能机制:①是依赖H+浓度或Ca++浓度的主动运输过程,这一过程需要消耗ATP,并且该转运方式在缺氧或添加代谢抑制剂的情况下被抑制②是谷胱甘肽(GSH)转运系统的转运方式及其类似转运,Vincerzini(1989[18])报道,GSH的跨膜转运与Na+、K+、LI+、Ca++、Mn++的浓度梯度有关,而与H+的浓度无关。目前对这两种机制的具体过程还不是非常清楚。由以上可见,小肽的转运不同于氨基酸,其转运过程是由多种因素共同参与作用的,因此小肽转运的具体过程还有待于更进一步的研究。
1.2 反刍动物寡肽的转运机制
与单胃动物不同的是反刍动物对小肽的吸收可分为肠系膜系统和非肠系膜系统(Webb,1993[19])。空肠、结肠、回肠、盲肠吸收的小肽进入肠系膜系统,而由瘤胃、网胃、瓣胃、皱胃、十二指肠吸收的小肽则进入非肠系膜系统,反刍动物的非肠系膜系统是小肽吸收的主要途径。反刍动物对小肽的吸收一种是以被动扩散形式进行的。Matthews(1991[10])用离体瘤胃上皮细胞和瓣胃上皮细胞研究小肽的吸收情况时发现,瘤胃上皮细胞和瓣胃上皮细胞是不饱和的被动扩散过程,并且瓣胃上皮细胞吸收小肽的能力强于瘤胃上皮细胞。反刍动物对小肽的另一种吸收形式是通过载体介导的主动运输过程。McCollum和Webb(1998[20])研究了羊瓣胃的Gly-Ser的吸收机制,结果表明,其转运也是由载体介导,依赖H+的浓度梯度进行的。
2 影响寡肽转运吸收的因素
2.1 肽链的性质
目前的研究认为,小肽比大肽、L型比D型、中性比酸、碱性肽更易吸收(钱利纯,1998[21])。二肽和三肽能完整的吸收,但三肽以上的寡肽是否能完整吸收还存在争议,Grimble等(1986[22])报道,肠道对于大于三肽的寡肽的吸收慢于二肽、三肽;Grimble和Silk(1989[23])指出,摄入肠道大于三肽以上的寡肽,在肠道内进一步水解为二、三肽后才能被动物吸收利用,这就降低了这些寡肽的吸收速度。而有研究者认为大于三肽的寡肽的吸收速度并不会逊色于二肽和三肽,但目前还未找到足够的证据来证明。Burston等(1972[24])报道,肽的氨基酸组成影响其吸收,当谷氨酸以谷氨酰胺赖氨酸形式而不以谷氨酰胺蛋氨酸形式供给时,大鼠小肠对其吸收速度加倍。另外,氨基酸残基构型也影响肽的吸收。当Lys位于N端与His构成二肽时,要比它位于C端时吸收速度快;而当它在C端与Glu构成二肽时,其吸收更为迅速。
2.2 日粮营养水平
对人体的研究发现,当限制饮食时肠道肽酶的活性下降,寡肽的释放量下降,而当供给蛋白质充足的日粮时,肽酶的活性则会升高,从而小肽的释放量增加。Webb等(1992[25])报道,长期对大鼠限饲,肠组织吸收L-Met和L-Met-L-Met的能力上升。Chen.G.等(1987[26])报道,黑白花奶牛日粮豆粕含量分别为14.5%,17.1%,20.6%时,其瘤胃吸收肽的量分别为22,33,34g/d,这说明奶牛对寡肽的吸收与其日粮的蛋白质水平有关。在消化过程中,寡肽的形成数量和比例与饲粮蛋白质品质有关。Savoie等(1987[27])对19种动物、植物性(豆科、谷物)蛋白质进行体外消化试验,在胃蛋白酶、胰蛋白酶的作用下,动物性蛋白质释放出的肽与游离氨基酸的比例高,豆科蛋白次之,而谷物蛋白质的释放量最低。乐国伟等(1996[28])对几种不同动植物性蛋白质饲料胃蛋白酶、胰蛋白酶水解产物的反相液相色谱分析表明,寡肽的释放量由大到小依次为酪蛋白、鱼粉、蚕蛹、豆粕、豆饼、菜籽饼、玉米蛋白粉,饲料蛋白质的寡肽释放量与有效氨基酸呈正相关。刘选珍等(1996[29])的试验表明,饲料寡肽释放量与碱性氨基酸含量相关的蛋白质的含量呈正相关。由这些试验可以看出,必需氨基酸含量高且平衡的优质蛋白在消化过程中易水解生成分子量低且数量多的寡肽,有利于寡肽的转运吸收;而必需氨基酸缺乏且不平衡的饲料蛋白质产生大量的游离氨基酸和少量的大分子量的肽片段,不利于寡肽的转运吸收(Meister,1987[30])。
2.3 加工、储存条件的影响
加工、储存条件是影响蛋白质消化过程中寡肽释放量的重要因素。Restani等(1992[31])在体外水解试验中发现,蒸制加工后的肉品与鲜肉相比,前者释放的寡肽量少,而冷冻干燥或鲜肉则释放较多的寡肽。Swaisgood和Catignari(1991[32])则指出,经过加热长期存放的豆粕,肽的释放量仅为有效Lys含量高的新鲜豆粕的63%,这可能是由于Lys的侧链-NH2因易发生Mallarld反应,使Lys残基与其毗邻的氨基酸肽健难以断裂,从而影响蛋白质的消化率。
2.4 动物方面的影响
由于单胃和反刍动物对小肽吸收的主要部位和转运系统的特点都不同,而且对以小肽和氨基酸两种方式沉积氮的比例也不同,因此不同的动物对寡肽的转运吸收是不同的。研究证明,动物吸收蛋白质是以小肽和游离氨基酸两种形式进行的,但单胃动物与反刍动物对蛋白质的吸收有着不同的特点。Gardner(1975[33])发现,小鼠小肠浆面的总氨基酸中约有20%是以小肽的形式存在。在反刍动物进入血液的总氨基澄中至少有50%是以小肽的形式被吸收的。Koeln和webb(1982[34])发现,在反刍动物门静脉中的氨基酸80%与小肽有关,只有20%的氨基酸以游离氨基酸的形式吸收,这说明反刍动物是以小肽吸收为主。因此,就两种动物的吸收部位来说,单胃动物吸收肽是在肠系膜系统,反刍动物吸收肽主要是在非肠系膜系统。由两系统吸收的小肽的数量见下表。Dirienzo(1990[35])以犊牛和绵羊为试验动物,对流经其肠系膜系统和非肠系膜系统的游离氨基酸和小肤进行定量研究,证实了上述结论。另外,动物所处的生理状态和代谢程度也会影响小肽的吸收。泌乳牛和绵羊在注射牛生长激素(BST)后,肌肉与乳房组织对小肽的利用程度加强(Mcdowell,1991[36]),这可能是由于BST等代谢调节剂加大了机体对与代谢变化有关的氨基酸的需要(Boyd等,1991[37])。
2.5 小肽载体(PepTl,PepT2)
小肽载体在小肠、肾对小肽的吸收中发挥着重要的作用(Shen等,2001[38]),肽载体对底物具有广泛的适应性,能够转运许许多多的化合物,包括小肽、肽类似物及非肽化合物,PepT1是低亲和力/高容量的肽载体,PepT2是高亲和力/低容量的肽载体。PepT1主要在消化道中表达,在肾脏中也有微弱的表达;PepT2主要在肾脏中表达(Fei等,1994[39]Liang等,1995[40];Miyamoto等,1996[41];Saito等,1996[42])。PepT2较PepTl对底物具有更高的亲合力(Terada等,2000[43]),但对底物表现出较严格地立体特异性,如对N末端含D一型氨基酸的耐受性比C末端为D一型的好.全D型的肽不能作为底物,对疏水性侧链体积大的底物,如含支链氨基酸、蛋氨酸、苯丙氨酸的肽具有较高的亲和力,而对亲水性、带电荷的小肽亲和力较小(Matthews,1991[10])。
2.6 其它因素
不同比例的小肽与游离氨基酸对动物氨基酸的吸收也有影响。小肽比例的增加能够显著提高氨基酸的吸收速度;提高游离氨基酸的浓度或比例时,并不能加快大多数氨基酸的吸收(施用辉等,1996[44])。日粮氮源也是影响肽吸收的因素。McCormick和Webb(1982[45])试验表明,饲喂干草、玉米和豆粕组成的传统日粮喂小牛,其血浆内循环肽氨基酸的浓度很高,肽氨基酸的浓度是游离氨基酸浓度的3倍多(683vs.191μmol/dl),并且能被后肢组织清除。而当饲喂纯化日粮如由尿素提供日粮氮时,血浆中循环肽氨基酸的浓度是游离氨基酸浓度的2倍多(407vs.178μmol/dl),但后肢组织几乎不清除。血浆内肽的清除依日粮氮源的不同而不同,以大豆作为氮源时血浆内肽能被后肢在、组织清除,而以尿素作氮源时后肢组织几乎不清除肽。这表明日粮氮源能够影响血浆内肽被组织利用的情况。