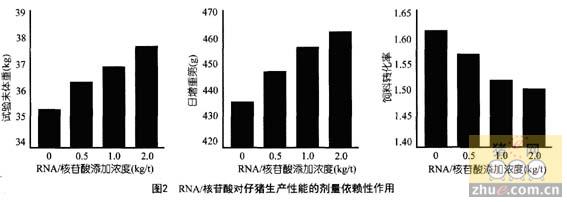
��״�����ἰ�������������е�Ӧ��
����: 2010-04-03 | ����: ��վ�Ѽ� | ��Դ: ��վԭ��
ժҪ���������ᣨcamp���ǽ��类���ֵ�ϸ������Ϣ���ݵĵڶ���ʹ����ϸ������Ҫ������������ʣ�����ϸ���ķֻ�����ֳ����л�����߷�Ӧ������Ҫ�ĵ������á�������Ҫ�����˻���������������ܼ��������������е�Ӧ�ã������ָ������������ҵ��Ӧ�ô��ڵ��й����⣬������Ӧ��ǰ������չ����
�ؼ��ʣ��������� �������� ��������
�������ᣨcamp����sutherland��1957�귢�֣���sutherland��1965�����camp��Ϊ���صĵڶ���ʹ���ۺ�[1]���������ǵĹ㷺�Ĺ�ע����Щ��������������ݷ�������ѧ���ۣ�������ҵ��ҽѧ������ѧ�ȷ����camp�����˹㷺���о�������camp�н鼤�ض�ϸ���������ֻ������Ѽ���ijЩ������ص���������ѧ���ܣ�һЩ�о��߰���Դ��campӦ�������������У�����camp����������������������ܺ�������Ʒ�ʵ����á�
1 ������������ɺͷֽ�
��ϸ���ܵ����̼�ʱ�������źŷ����������������γɸ����壬Ȼ��ϸ��Ĥ�ϵ�gs-���ף��������gs-�����ټ���ϸ��Ĥ�ϵ������ỷ��ø��ac������atp��ȥһ�������������camp�������˵������źż����壨h����ϸ��Ĥ�ϵ����壨rs������γ����帴���hrs������������Ĺ������ı䣬��¶��������gs��ϵ�λ�㣬����hrs��gs��ϣ��γ�hrsgs�����壬�Ӷ�����gs���ף�gs�ϵ�gdp��gtpȡ����ͬʱ�ͷ�gdp�ͦ¦��ǻ������Ǧ��ǻ���¦ø���������룬ͬʱ��¶�����ǻ���ac�Ľ��λ�㣬ʹ���ǻ���ac��϶�����ac���������ac����atp����camp���ǻ���gtpø����ʹ��ϵ�gtpˮ��Ϊgdp��ͬʱ���ǻ��ָ����������ac���룬�Ӷ�acʧ������ǻ�������¦��ǻ���ϡ����ɵ�camp��Ϊ�ڶ���ʹͨ������apk��camp�����Ե���ø����ʹ��ϸ���������ữ���Ӷ�����ϸ����Ӧ��camp�����ֱ��������ø��pde��ˮ���5 -amp��ʧ�camp���ɺͷֽ��������mg2+�Ĵ��ڡ�ac��pde���Դ��ڶ�����ͬ�������ϸ���ڵ� camp��Ũ�ȣ��Ӷ�Ӱ��ϸ������֯�����ٵĹ��ܡ���ac�Ļ�������ʱ��camp��Ũ�����ߣ���pde��Ũ������ʱ��camp��Ũ�Ƚ��͡�pde��camp�ĵ��أ�����ȡ����pde�Ļ���������أ���ȡ����ϸ����pde����ɡ���ϸ���ֲ����й�pde���ڿ���ϸ����ˮƽ������о��ѳ�Ϊ�������о���һ���ȵ�[2]��
2 ���������ڶ������ڵĺ������ֲ�
��1957��sutherland�����ڸ����Ƚ��з���camp֮������½���ںܶ���֯�������Ρ�������״������֧���ܡ��Դ��塢ѪС�塢��֭��غ�衢�������֯����Һ�з�����camp���ڡ����鶯�����ϸ���⣬������֯�ж��зֲ���ͨ����ͬ��ϸ���У�cgmp��camp��ˮƽ�͵Ķࡣ[3]��������£�ϸ���ڵ�camp��Ũ��Ϊ0.1~1um�����ڼ��ػ�Ӧ���������£������ߵ�һ�ٱ����ϡ�Ѫ���е�camp��ϸ���еĵͣ�ԼΪ10nm�����ڵ�camp�����ͷ���Ѫ��˵��Ѫ��camp�ı仯����һ���ij̶��Ϸ�ӳ�������ٵ������仯��
3 �����������������
camp������ЧӦ�Ƕ��ģ���ͬ��ϸ����camp�ķ�Ӧ��ͬ������Ҫȡ����ϸ���е�øϵ�����ࡢ�����Լ��������ܵ������á�
3.1 ���������ϸ����л�ĵ���
camp����ϸ���������л������ͨ������ø�Ļ�����ʵ�ֵġ�����atp���ڵ������£�apk���Դ�ϸ���������л�ؼ�ø���Եļ�������ữøb��֬����֯֬��ø����ijЩø�����ƣ����л��Ե���ԭ�ϳ�ø���������յ���ijЩ��л��Ӧ�ļ��ٻ����ơ�[4]
1962��krebs�����о���camp����ԭ�ϳɺ���ԭ�ֽ�øϵ�ĵ��ڡ��������ؼ��ȸ�Ѫ���صȼ��ؿ�ʹcampˮƽ���ߣ�����apk��apk����������ԭ���ữ��ø��ʹ��ԭ���ữø���ữ���Ӷ���ԭ���ữø�����Ե�b��ʽת��Ϊ�л��Ե�a����ʽ�����ߴ���ԭ�ֽ�Ϊ1-p-�����ǡ�camp��ͨ��apkʹ��ԭ�ϳ�ø���ữ�����¾��л��Ե���ԭ�ϳ�øת��Ϊ���Ե���ԭ�ϳ�ød���Ӷ�������ԭ�ĺϳɡ�camp���ܼ����ǽͽ��е�һ���ؼ�ø??������Ǽ�ø����6-�����������1��6-��������ǡ����⣬camp������ֹatp��������ǵ����ơ�
���⣬camp����ͨ��apk����֬������ø��ʹ֬��ˮ��ؼ�ø??֬��ø���ữ������Ӷ��ٽ�֬��ˮ��Ϊ���ͺ������֬���ᡣ֬���ᱻת�Ƶ�ѪҺ�У������Ѫ������ϣ�Ȼ��ת�˵�������֯�У��ر������ࡢ���⡢������֯������������������ѭ��������atp����Ϊϸ������Դ�����磬camp����̼��������ø�����߿�ʹ̼����ø���ữ�������co2�γ�̼�ᣬ̼���ٷֽ�ų�h+���Ե���ϸ�������ƽ������Ҫ�����ã����Ҵ˹��̿�ʹθ�����࣬������������[4,5]
3.2 ��������Լ��غϳɺͷ��ڵ�Ӱ��
camp���е������ʵĺϳɣ��ٽ����ص�����[6]������֤�ݱ�����һЩ�����ټ��شٽ��μ����غϳ���ͨ��camp;�����ڵġ���������Ƥ�ʼ��ؽ�ϵ�������Ƥ��ϸ������ac������camp��Ũ�ȣ�����pka���������ữ����Ƥ��ͪ��ȩ��ͪ�ĺϳ�ø�����ѳ�ϸ���У�Ҳ�����Ƶ�����������ݼ���ͨ��camp;�����Ӵƶ�������ͪ�ĺϳ�[5]. camp���յ�gh���ͷţ��Ӷ��ٽ������ڵ����ʡ�dna��rna�ĺϳɣ����ܼ�ǿ֬���ֽ⣬�̼����嵰���ʵĺϳɡ�[7��8]
3.3 ������������߹��ܵ�Ӱ��
�ݱ������Դ���Ƣ���ܰ�ϸ���������������������м���camp�����ʣ�һ��ʱ�����Ƣ���ܰ�ϸ��������tϸ���Ŀ�ԭ��Ӧ���ȶ������b�ܰ�ϸ���������ӣ����γɿ���ˮƽ���ӡ����͵ȣ�2001����̽�ּ��������ܲ�ԭ�����Ⱦʱ��ϸ����ʹ���ʶ����߹��ܵĵ�������ʱ���֣��Դ�Ⱦ�Է����Ҳ�����(ibdv)ǿ���깥�����⼦����?����������ǿ�����������ibdv�����������߷�Ӧ��Ѫ��camp������������ʾ����?����������ǿʹcamp�����뿹ibdv����������ϸ����ЧӦ����ӵĺϳ��й�[9]��
3.4 ���������Ĥ�����Եĵ���
camp�ɴ�ʹ����ϸ��Ĥ��ijЩ�������ữ��ʹ�乹�ͷ����ı䣬�Ӷ�����Ĥ��һЩ���ʵ�ͨ�ԡ����������١��ȸ�Ѫ������֬��ϸ��Ĥ�ϵ���Ӧ�����Ϻ��ϳ�camp��camp��ʹ��ϸ��Ĥ�ϵ������ữ������ı䣬�����ǵ���ת�������ǵ������������½���������֤�ݱ������ɵ��°��������յ��½����ں�ϸ���У�camp����ϸ��Ĥ�ϵĵ���ø��ʹĤ�ϵ�spectin�������ữ�Ժ�ϸ��Ĥ���������ʼ���ϸ������̬������Ϊ��Ҫ�ĵ������á���ѪС���У�camp��ͨ��apk��Ч�ش̼�Ĥ�ϵ�һ�ַ�����Ϊ22000�ĵ������ữ����ͨ���Ը������Ӱ�죬����ѪС��ۼ��������ȹ��ܡ����ļ�ϸ���У�camp��ʹ�ļ�ϸ����ͨ���������ữ��ʹĤ��ca2+��ͨ����ǿ������ca2+�������Ӷ�ʹ�ļ����������ӣ����ʼӿ졣�������������֤����ѹ�ص�ͨ��camp��ʹϸ��Ĥ�������ʸı䣬���Ӷ�ˮ�����ա�[4��5]
3.5 �����������ϸ��������
mcafee��1971������֤��camp������ͻ�����ݡ�Ŀǰ��Ϊ����ijЩ��ϸ���˷�ʱͻ��ǰ��ĩ���ͷŵ��ʡ�������ͻ����Ĥ����Ӧ�����岢����ac����ͻ����Ĥ�ϳ�camp����������pka��ͨ��Ĥ�������ữ�ı�Ĥ�����ӵ�ͨ�ԣ��Ӷ�Ӱ����ϸ�����˷��ԡ�
����֯�ں��и�ˮƽ��camp�����л����ø�����ԡ����衢�Լ�Һ���������ж��д�����camp���ڡ��ڼ��������У�camp������ߣ��ȷ�����֯Լ��10����ac��camp�������ø����Ҳ��������֯��10?20��������˵��������֯�е�camp�ĺϳɺͷֽ��ٶ�ԶԶ����������֯��Ҳ˵��camp������֯��������Ҫ�����á�[5]
3.6 ��������Ի������ĵ���
camp��һ����Ҫ�Ļ������ĵ�������[10]����ԭ��������camp����Ϊ��ֱ�ӻrna�ۺ�ø�Դٽ�ת¼����ͨ����ø�ħ����ӵ����ữ��ʵ�ִٽ�mrnaת¼��
���������о����������ϸ���е�camp��������ת¼���ӵ����йء�montminy�ȣ�1986����������camp�յ�ת¼����˻������������Χ�ຬ��һ�»����һ�µ�8������ԵĻ�������5��-tgacgtca-3��������֮Ϊcamp��Ӧ���У�camp-responsive element��cre��������Щ����ʶ��camp�źŵ���Ҫ��λ��ͬʱ�����ǻ�����camp�յ��İл�����ﻹ��Ҫpka�ļ��campˮƽ������pka��pka�ֿ���ͨ��ʹijЩ�����ת¼���ӽ������ữ���鵼camp����Ļ������[11]��1987�꣬montminy��֤ʵ����һ�Ʋ⡣[12]����ʵ�������pka��ʹ�鵰�����ữ�����ữ���鵰�����ڴ���״̬������ĸı䣬��dna����ɳڶ����룬�Ӷ�������鵰����λ�������ƣ�ʹת¼���Խ��С�������ʵ�鷢�֣�������pka��ʹ���鵰�����ữ�����ữ�����Ե���������ǿ�����н϶�ĸ���ɣ��������ɵ��鵰���н�ǿ�����������ϣ�ʹ�鵰����dna���룬����鵰��dna�����ֶ�����ת¼��[4,5]
3.7 ���������ϸ����ֳ��ֻ��ĵ���
ϸ������ֳ��ֻ���ϸ��������������������ֳ����ϸ����������dna���ƺ�ϸ�����ѣ���ͨ��ϸ��������ʵ�ֵġ��ֻ���ζ��ϸ���ڵ�dnaͨ��ת¼����mrna���ٽ�һ���ϳ�רһ�Եĵ����ʣ�ø�������յ���ϸ����̬���ṹ��������ɺ��ܵȸ���ˮƽ�ϵIJ��졣ϸ����ֳ��ֻ���һ�Լ�����ϵ����ì�ܵĹ��̣�camp�ڵ������ì����������Ҫ������[13,14] ��camp������ϸ��������ϸ�����ѡ��ٽ��ֻ������á���ˣ�����ʹϸ����camp�������ߵ����ؾ��ܽ���ϸ���������ٶȣ�����ϸ������ֳ�����ٽ�ϸ���ķֻ���[4,5]��������Ϊcamp��ϸ����ֳ����˫��ЧӦ������g0����g1��ʱ��ϸ����ֳ���дٽ����ã�������g1��ʱ�����������á�[15]
���⣬camp��������һ���Ĺ�ϵ����ϸ��������ϸ���е�camp�������в���ģ�������ϸ���ڵ�campһ���������ϸ��ˮƽ��ϸ�����ٻָ�����ϸ��ʱ�����ڵ�campˮƽҲ�漴�ָ�������[4]
4 ��������������ҵ�е�Ӧ��
camp����߶�����������ܺ������ƷƷ�ʡ������û���Ϊ��camp��Ϊ���صġ��ڶ���ʹ�������ø��ʹ��лø������ǿ���Ӷ���ǿ���ڵ��ĺϳɣ����춯��������ٶȣ��յ����أ����������صȣ���ø�ĺϳɣ��ٽ�����ĺϳɴ�л��
4.1 ���������ڼ��������ϵ�Ӧ��
4.1.1 �������������������ϵ�Ӧ��
����������������camp���������õ��������ܺ�����Ʒ�ʵ�Ӱ����һ�¡���ˮƽ��camp������֬����֯��hsl���ʹ֬���ֽ�������ǿ��[16]ͬʱcamp���ɼ����ȵ�����֬��ϸ���Ľ��[17]���ԶԿ��ȵ��ض���֬�����Ĵٽ����á�������ȣ�1992������Ƥ��ע��camp��pde�����Ƽ������������������������������ɣ�����������ʡ��ۼ�����ͺ��˱ȣ����һ�ʹѪ����֬�ʺ�����֬��ø�Ļ������ߡ���ʾ��֬���ͺ�Ѫ֬���߿�����camp������ʹ����֬��ø�������ߡ�������֬��Ա�йأ�����֬�Ĵ����ֽ����������ڽ��͵����ʵķֽ⣬�Ӷ���ǿ�����������ڵij���[18]��
camp���������Ҳ�����ԵĴ��������á���ʿ����1997�����������������������ע��camp�Ʊ�Ʒ�ͱ�Ʒ�Ƽ������������������á�[19]�����µȣ�1993��ע��camp���������������Ե�����Ч����������ʹ��ʱ����ӳ�����Ч�������½���[20]
���⣬camp�����������IJ�ë����ë��ά���ȣ��ı���ë�и��ְ�����ĺ���������ë�����Ļ���������ͨ��������ë�������ڻ�����ë�ҵ�Ӫ����Ӧ��ʵ�ֵġ�[21]
4.1.2 �������������������ϵ�Ӧ��
����ʮ����������ͨ���Դ���С�������ú�������ţ�����ȶ������ٺ�����camp���о�������camp����������������������Ҫ���á����﷼�ȣ�1993�������˲�ͬ�����ɽ�����л�������ı仯������ǰ��������������ߣ�����ά���ڣ������15�죬���е�campŨ��һֱά���ڽϸ�ˮƽ��������߷��ڣ�camp�������ߣ���������ڣ�ֹͣ����ǰ1�������ң���ֻ��������������½������е�camp������½�������Ϊcamp��Ҫ�ٽ���֭���ڣ�cgmp��Ҫ�Ǵٽ���֭���ɣ����߳�һ�������仯��[22]�����ܵȣ�1995����ͬƷ��ţ��Ѫ��campˮƽ����Ϊ������ͬ�ı仯���ɡ�����ǰ��һ�ܳ�С�IJ�����Ȼ��Ѹ���������ڷ����45��ﵽ��ֵ�������90��Ѫ��camp���½������ơ� [23]³��̫�ȣ�1994�����������е�camp�ı仯�������еı仯�෴[24]��shiffield ����ע����Ҷ��أ�camp�ٽ������ķ����ɹ���ʹ��ֲ���������ڵ�ţ������֯�����ͷ���[25]�����о���camp�������������������ɽ������������������ȣ�2002����������һ����Ũ�ȷ�Χ�ڣ�camp������������ɽ��������Ƥϸ����ֳ�дٽ����ã����ʼ����������ԣ���campŨ�ȹ���ʱ�����������á�camp�Ը�ϸ���������ߵ�Ӱ�����ڶ��������ڡ���Դ��camp�ܴٽ�����������ɽ��������Ƥϸ��������֭��ɽ���ҵ��ı�� rillema���������ص�����������camp��ˮƽ��һ��������ԡ�[26]Ȼ��camp����ε�������ϸ�������顢�����ģ�Ŀǰ�в�ʮ��������������������camp�ٽ�����ǰҶgh��prl(����̼���)�����鼤�ص����ɺͷ��ڣ��ӿ���弸��������֯ϸ���Ĵ�л�����ȴ���ƶ��ֶ�����������ϸ��dna��rna�����ǡ��鵰����֬�ĺϳ�[27,28]������������Ϊ��camp�ɽ��ʹ���С�������õ����ٺ��Ǻ��գ�rna�������ǡ���֬���ҵ��ȵĺϳɣ�����һЩø�Ļ��ԣ��������������ɡ�[29,30]������Ϊcamp���������������أ�camp�ٽ���֬�ϳɣ������鵰�ϳɡ�[31]
4.2 ���������ڼ��������е�Ӧ��
��Դ��camp�������ٽ���ά�����м�����������ؼ��ʡ��ȼ��ʡ�����֬�ʣ�������Щ��Ӧ���ż��������Ӷ�����[32]�������ỹ���⼦��л��Ӱ�죬���ɽ������е�֬���������ٽ������ʻ��ۣ��Ը�����֯Ҳ�����Ƶ�Ч��������ʹѪ���ܵ�����֬��������[33]��camp������֬����л�����úͲ����ද��������ͬ��camp�ܴٽ������ද��ļ�������֬ø��hsl���������ߣ��ٽ�����֬���ķֽ��Ѫ֬�ĺ������ߣ��Ӷ��������е�֬����������������Ʒ�ʡ����ڼ����У������������ض����ȸ�Ѫ���أ�������Ҫ��֬�⼤�ء�ţ����ȣ�1998��������Դ�Ի���������⼦���ڵ�gh�ķ��ڡ���л��һ���Ĵٽ����ã������������������Կ��ڶ����������£�������ȴ���ڶ����飬�������camp���⼦����Ѫ��ϵͳ���е��������йأ���һ������Ѫ���е����ʺ����������ڶ����飬�������������й�[34]�����⣬camp����������ߵ����IJ������ܣ����Ƶ��ǵ�Ʒ�ʣ��ɴ��������ľ���Ч��[35]��
5 �йش��ڵ�������Ӧ��ǰ��չ��
Ŀǰ�й�camp����������ϵͳ�о��Ƚ��٣�����Ŀǰ�й�camp���о����������ע��ķ����������ɸ����ģ���ƹ�Ӧ�ô��������ѡ����⣬������Դ��campȷ�����û���������Դ��camp�к�Ӱ��������д��ڽ�һ���о���
��Ȼ���й�camp���о���������ҵ�ϵ�Ӧ�ô����������⣬����Ϊ���������ĵ��ڼ���camp������Դ�㣬��ȡ���Ʊ�������ŵ㡣���⣬�����������������ǵ��������Ʒ�в������������������������쳣��Ŀǰ�ѱ����á���ѡ�ü��صĵڶ���ʹ�����Ӽ��ص�ЧӦ���ȿ�����߶����������ܺ������ƷƷ�ʣ������κβ���ЧӦ����ˣ������camp���о����������������е�Ӧ�þ������õ�ǰ����
�ο����ף�references��
[1] sutherland e m, ye i & butcher r w. the action of epinephrine and the role of adenyl cyclase in hormone action [j]. recent progress in hormone research, 1965,21,623~646
[2] ������. ϸ����Ϣ�����[m]. ����������ҽ�ƴ�ѧ���й�Э��ҽ�ƴ�ѧ���ϳ����磬1998��168~176
[3] ����ɭ����Ԭ־��.�����������������ڴ�л���ɵ��о�[j]. ������ҽѧ����2000��31��2����124~130
[4] ����. camp��ϸ����л�еĵ�������[j].���ﻯѧ������������չ��1981��3��11~14
[5] ���ҵ�������֣�������. ϸ���ź�ת��[m] (�ڶ���). ��������ѧ���������磬1997��52~70
[6] gerosa ma. cyclic nucleotide deesin meduloblastomas correlative study of tumoral and cerebrospinal fluid levers [j]. neurosurgery, 1980, 7:359
[7] dana s, et al. induction of human growth hormone promoter activity by the adenosine 3��,5��-monophsphate pathway involves a noval responsive element [j]. ca., 1989,17(3): 72~75
[8] ganced j m, adelstein r s, morgan k g et al. regulations by cyclic nucleotides and phorbol ester of calcium efflux form cultured bovine adrenal chramaffincells [j]. tibs, 1985,10:5
[9] ���ͣ�ţ����ʯ���۵�. �����������⼦�ڷ��ڼ����߹��ܵĹ�ϵ[j]. �й���ҽѧ����2001��21��3����278~280
[10] monall e. expression of myosin and action in e. coli. merhods, enzymot, 1991, 196:368~389
[11] montminy m r, sevatino k a, wagner j a, et al. identification of a cyclic amp-responsive element within in rat somatostation gene [j]. proc natl acad sci usa, 1986,83:6682~6686
[12] montminy m r, bilezkjian l m. binding of a nuclear protein to the cyclic-amp element of the somatostation gene [j]. nature, 1987,328:175~178
[13] burgering bmt, bos jl. regulation of ras-mediated signing: more than one way to skin a cat[j]. trends biochem sci, 1995a,20:18~22
[14] dumont je, jauniaux jc, roger pp. the cyclic amp-mediated stimulation of cell proliferation [j]. trends biochem sci, 1989,14:67~71
[15] miyazaki m, wahid s, bai l et al. effects of intracellular cyclic amp and cyclic gmp levels on dna synthesis of young-adult rat hepatocytes in primary culture [j]. exp cell res, 1992, 200: 404~409
[16] ecan j j, greenberg a s, chang m k et al. mechanism of hormone-stimulated lipolysis in adipocyles: translocation of hormone-sensitive lipase to the lipid storage droplet[j]. proc. natl. acad. sci. usa,1992,89:8537~8541
[17] liu c y and mills s e. decreased insulin binding to porcine adipocytes in vitro by beta-adrenergic agonists[j]. j sci, 1990,68:1603~1609
[18] �����壬�ν�����³��̫����Դ����Դ��camp����������֬�ʺ͵����ʳ�����Ӱ�죮����ũҵѧ����1992��1(4)��19?22
[19] ��ʿ��. ��״����������������Ч�����о�[j]. ����������ҽ��1997��6��14~16
[20] �����£���ʿ������֮����. ��״�������������������Ч����̽[j]. �ຣ������ҽ��־��1993��23��3����23~24
[21] black j l, reis p j, ed. physiological and environmental limitation to wool growth[j]. armidale; university of new england publishing unit,1979,337~352
[22] ���﷼��ŷ�����죬��־��. ��ͬ�����ɽ�����л�������ı仯[j]. ����ũҵѧ����1993��2��4����81~84
[23] �����ܣ��ž��䣬�����ٵ�. ��ţѪ����������ˮƽ��������ܵĹ�ϵ[j]. ����ũҵ��ѧѧ����1995��17��4����60~62
[24] ³��̫���ξ��ޣ�������. ��������������[j]. �й���ţ��1994��6��38~40
[25] sheffield lg.. influence of cholera toxin (an adenylafe cyclase activator ) on deoxyribonucleic synthesis of bovine mammary tissue in vitro and in athymic nude mice[j]. j dairy sci. 1989,72(1):41~48
[26] rillema j a. cyclic nucleotides and the effect of prolactin on uridine incorporation into rna in mammary gland explants of mice[j]. horm. metab res, 1975,7:45
[27] clayto r n. cyclic adenosine nucleotides and growth hormone-releasing factor increase cytosolic growth hormone messenger rna levers in cultural rat pituitary cells[j]. j edocr, 1986,110:51~53
[28] mature r a. transcriptional regulation of prolation gene by ergocription and cyclic amp[j]. nature, 1981,294:94~97
[29] perry j w, oka t. cyclic amp as a negative regulator of hormonally induced lacto genesis in mouse mammary gland organ culture[j]. prot natl. acad. sci usa,1980,77:2093~2097
[30] sheffield l g. cyclic nucleotides concentrations and protein kinase activities of bovine mammary tissues maintained in athymic nude mice: effects of mammogenic and lactogenic hormone[j]. j. dairy sci, 1988,71:75~83
[31] �ԺƱ�³��̫. �������������ָ����������ܹ�ϵ���о�[j]. ����ũҵ��ѧ��1992��2��21~22
[32] �Ÿ�÷��Ѧ���������. ��Դ�Ի���������⼦�������ܵ�Ӱ��[j].�й�������־��2001��37��3����25~26
[33] �Ÿ�÷�������ƣ����µ�. ����������⼦ѪҺ����֯����ָ���Ӱ��[j]. �й����ݣ�2000��22��12����10~11
[34] ţ���ᣬ�������������ĵ�. ��Դ�Ի��������aa�⼦����������Ӱ��[j].�й���ҽѧ����1998��18��3����299~301
[35] ţ���ᣬ�����ѣ��³�����. ��Դ��camp�Ժ�������Ʒ�����������ܵ�Ӱ��[j].��������ҽ��2000��32��5����17~18
camp and its application in animal science
wang shanfeng��chen anguo
��animal science college, zhejiang university��
abstract as an important intracellular second messenger in cells, cyclic adenosine monophosphate is involved in cellular differentiation, cellular proliferation, cell metabolism and immunoreactions. moreover��it plays an important role in the above process. in the article, physiological function of camp and its application in livestock husbandry are reviewed. finally, the correlative problem and the prospect of application of camp in animal science are mentioned.
keywords: camp, physiological function, animal science.
����飺
���Ʒ棬�У���1976?�����㽭��ѧ�����ѧѧԺ2001��˶ʿ�о���
�°��������ڣ�˶ʿ�о�����ʦ��ͨѶ��ַ�뵥λͬ��һ����
�ؼ��ʣ��������� �������� ��������
�������ᣨcamp����sutherland��1957�귢�֣���sutherland��1965�����camp��Ϊ���صĵڶ���ʹ���ۺ�[1]���������ǵĹ㷺�Ĺ�ע����Щ��������������ݷ�������ѧ���ۣ�������ҵ��ҽѧ������ѧ�ȷ����camp�����˹㷺���о�������camp�н鼤�ض�ϸ���������ֻ������Ѽ���ijЩ������ص���������ѧ���ܣ�һЩ�о��߰���Դ��campӦ�������������У�����camp����������������������ܺ�������Ʒ�ʵ����á�
1 ������������ɺͷֽ�
��ϸ���ܵ����̼�ʱ�������źŷ����������������γɸ����壬Ȼ��ϸ��Ĥ�ϵ�gs-���ף��������gs-�����ټ���ϸ��Ĥ�ϵ������ỷ��ø��ac������atp��ȥһ�������������camp�������˵������źż����壨h����ϸ��Ĥ�ϵ����壨rs������γ����帴���hrs������������Ĺ������ı䣬��¶��������gs��ϵ�λ�㣬����hrs��gs��ϣ��γ�hrsgs�����壬�Ӷ�����gs���ף�gs�ϵ�gdp��gtpȡ����ͬʱ�ͷ�gdp�ͦ¦��ǻ������Ǧ��ǻ���¦ø���������룬ͬʱ��¶�����ǻ���ac�Ľ��λ�㣬ʹ���ǻ���ac��϶�����ac���������ac����atp����camp���ǻ���gtpø����ʹ��ϵ�gtpˮ��Ϊgdp��ͬʱ���ǻ��ָ����������ac���룬�Ӷ�acʧ������ǻ�������¦��ǻ���ϡ����ɵ�camp��Ϊ�ڶ���ʹͨ������apk��camp�����Ե���ø����ʹ��ϸ���������ữ���Ӷ�����ϸ����Ӧ��camp�����ֱ��������ø��pde��ˮ���5 -amp��ʧ�camp���ɺͷֽ��������mg2+�Ĵ��ڡ�ac��pde���Դ��ڶ�����ͬ�������ϸ���ڵ� camp��Ũ�ȣ��Ӷ�Ӱ��ϸ������֯�����ٵĹ��ܡ���ac�Ļ�������ʱ��camp��Ũ�����ߣ���pde��Ũ������ʱ��camp��Ũ�Ƚ��͡�pde��camp�ĵ��أ�����ȡ����pde�Ļ���������أ���ȡ����ϸ����pde����ɡ���ϸ���ֲ����й�pde���ڿ���ϸ����ˮƽ������о��ѳ�Ϊ�������о���һ���ȵ�[2]��
2 ���������ڶ������ڵĺ������ֲ�
��1957��sutherland�����ڸ����Ƚ��з���camp֮������½���ںܶ���֯�������Ρ�������״������֧���ܡ��Դ��塢ѪС�塢��֭��غ�衢�������֯����Һ�з�����camp���ڡ����鶯�����ϸ���⣬������֯�ж��зֲ���ͨ����ͬ��ϸ���У�cgmp��camp��ˮƽ�͵Ķࡣ[3]��������£�ϸ���ڵ�camp��Ũ��Ϊ0.1~1um�����ڼ��ػ�Ӧ���������£������ߵ�һ�ٱ����ϡ�Ѫ���е�camp��ϸ���еĵͣ�ԼΪ10nm�����ڵ�camp�����ͷ���Ѫ��˵��Ѫ��camp�ı仯����һ���ij̶��Ϸ�ӳ�������ٵ������仯��
3 �����������������
camp������ЧӦ�Ƕ��ģ���ͬ��ϸ����camp�ķ�Ӧ��ͬ������Ҫȡ����ϸ���е�øϵ�����ࡢ�����Լ��������ܵ������á�
3.1 ���������ϸ����л�ĵ���
camp����ϸ���������л������ͨ������ø�Ļ�����ʵ�ֵġ�����atp���ڵ������£�apk���Դ�ϸ���������л�ؼ�ø���Եļ�������ữøb��֬����֯֬��ø����ijЩø�����ƣ����л��Ե���ԭ�ϳ�ø���������յ���ijЩ��л��Ӧ�ļ��ٻ����ơ�[4]
1962��krebs�����о���camp����ԭ�ϳɺ���ԭ�ֽ�øϵ�ĵ��ڡ��������ؼ��ȸ�Ѫ���صȼ��ؿ�ʹcampˮƽ���ߣ�����apk��apk����������ԭ���ữ��ø��ʹ��ԭ���ữø���ữ���Ӷ���ԭ���ữø�����Ե�b��ʽת��Ϊ�л��Ե�a����ʽ�����ߴ���ԭ�ֽ�Ϊ1-p-�����ǡ�camp��ͨ��apkʹ��ԭ�ϳ�ø���ữ�����¾��л��Ե���ԭ�ϳ�øת��Ϊ���Ե���ԭ�ϳ�ød���Ӷ�������ԭ�ĺϳɡ�camp���ܼ����ǽͽ��е�һ���ؼ�ø??������Ǽ�ø����6-�����������1��6-��������ǡ����⣬camp������ֹatp��������ǵ����ơ�
���⣬camp����ͨ��apk����֬������ø��ʹ֬��ˮ��ؼ�ø??֬��ø���ữ������Ӷ��ٽ�֬��ˮ��Ϊ���ͺ������֬���ᡣ֬���ᱻת�Ƶ�ѪҺ�У������Ѫ������ϣ�Ȼ��ת�˵�������֯�У��ر������ࡢ���⡢������֯������������������ѭ��������atp����Ϊϸ������Դ�����磬camp����̼��������ø�����߿�ʹ̼����ø���ữ�������co2�γ�̼�ᣬ̼���ٷֽ�ų�h+���Ե���ϸ�������ƽ������Ҫ�����ã����Ҵ˹��̿�ʹθ�����࣬������������[4,5]
3.2 ��������Լ��غϳɺͷ��ڵ�Ӱ��
camp���е������ʵĺϳɣ��ٽ����ص�����[6]������֤�ݱ�����һЩ�����ټ��شٽ��μ����غϳ���ͨ��camp;�����ڵġ���������Ƥ�ʼ��ؽ�ϵ�������Ƥ��ϸ������ac������camp��Ũ�ȣ�����pka���������ữ����Ƥ��ͪ��ȩ��ͪ�ĺϳ�ø�����ѳ�ϸ���У�Ҳ�����Ƶ�����������ݼ���ͨ��camp;�����Ӵƶ�������ͪ�ĺϳ�[5]. camp���յ�gh���ͷţ��Ӷ��ٽ������ڵ����ʡ�dna��rna�ĺϳɣ����ܼ�ǿ֬���ֽ⣬�̼����嵰���ʵĺϳɡ�[7��8]
3.3 ������������߹��ܵ�Ӱ��
�ݱ������Դ���Ƣ���ܰ�ϸ���������������������м���camp�����ʣ�һ��ʱ�����Ƣ���ܰ�ϸ��������tϸ���Ŀ�ԭ��Ӧ���ȶ������b�ܰ�ϸ���������ӣ����γɿ���ˮƽ���ӡ����͵ȣ�2001����̽�ּ��������ܲ�ԭ�����Ⱦʱ��ϸ����ʹ���ʶ����߹��ܵĵ�������ʱ���֣��Դ�Ⱦ�Է����Ҳ�����(ibdv)ǿ���깥�����⼦����?����������ǿ�����������ibdv�����������߷�Ӧ��Ѫ��camp������������ʾ����?����������ǿʹcamp�����뿹ibdv����������ϸ����ЧӦ����ӵĺϳ��й�[9]��
3.4 ���������Ĥ�����Եĵ���
camp�ɴ�ʹ����ϸ��Ĥ��ijЩ�������ữ��ʹ�乹�ͷ����ı䣬�Ӷ�����Ĥ��һЩ���ʵ�ͨ�ԡ����������١��ȸ�Ѫ������֬��ϸ��Ĥ�ϵ���Ӧ�����Ϻ��ϳ�camp��camp��ʹ��ϸ��Ĥ�ϵ������ữ������ı䣬�����ǵ���ת�������ǵ������������½���������֤�ݱ������ɵ��°��������յ��½����ں�ϸ���У�camp����ϸ��Ĥ�ϵĵ���ø��ʹĤ�ϵ�spectin�������ữ�Ժ�ϸ��Ĥ���������ʼ���ϸ������̬������Ϊ��Ҫ�ĵ������á���ѪС���У�camp��ͨ��apk��Ч�ش̼�Ĥ�ϵ�һ�ַ�����Ϊ22000�ĵ������ữ����ͨ���Ը������Ӱ�죬����ѪС��ۼ��������ȹ��ܡ����ļ�ϸ���У�camp��ʹ�ļ�ϸ����ͨ���������ữ��ʹĤ��ca2+��ͨ����ǿ������ca2+�������Ӷ�ʹ�ļ����������ӣ����ʼӿ졣�������������֤����ѹ�ص�ͨ��camp��ʹϸ��Ĥ�������ʸı䣬���Ӷ�ˮ�����ա�[4��5]
3.5 �����������ϸ��������
mcafee��1971������֤��camp������ͻ�����ݡ�Ŀǰ��Ϊ����ijЩ��ϸ���˷�ʱͻ��ǰ��ĩ���ͷŵ��ʡ�������ͻ����Ĥ����Ӧ�����岢����ac����ͻ����Ĥ�ϳ�camp����������pka��ͨ��Ĥ�������ữ�ı�Ĥ�����ӵ�ͨ�ԣ��Ӷ�Ӱ����ϸ�����˷��ԡ�
����֯�ں��и�ˮƽ��camp�����л����ø�����ԡ����衢�Լ�Һ���������ж��д�����camp���ڡ��ڼ��������У�camp������ߣ��ȷ�����֯Լ��10����ac��camp�������ø����Ҳ��������֯��10?20��������˵��������֯�е�camp�ĺϳɺͷֽ��ٶ�ԶԶ����������֯��Ҳ˵��camp������֯��������Ҫ�����á�[5]
3.6 ��������Ի������ĵ���
camp��һ����Ҫ�Ļ������ĵ�������[10]����ԭ��������camp����Ϊ��ֱ�ӻrna�ۺ�ø�Դٽ�ת¼����ͨ����ø�ħ����ӵ����ữ��ʵ�ִٽ�mrnaת¼��
���������о����������ϸ���е�camp��������ת¼���ӵ����йء�montminy�ȣ�1986����������camp�յ�ת¼����˻������������Χ�ຬ��һ�»����һ�µ�8������ԵĻ�������5��-tgacgtca-3��������֮Ϊcamp��Ӧ���У�camp-responsive element��cre��������Щ����ʶ��camp�źŵ���Ҫ��λ��ͬʱ�����ǻ�����camp�յ��İл�����ﻹ��Ҫpka�ļ��campˮƽ������pka��pka�ֿ���ͨ��ʹijЩ�����ת¼���ӽ������ữ���鵼camp����Ļ������[11]��1987�꣬montminy��֤ʵ����һ�Ʋ⡣[12]����ʵ�������pka��ʹ�鵰�����ữ�����ữ���鵰�����ڴ���״̬������ĸı䣬��dna����ɳڶ����룬�Ӷ�������鵰����λ�������ƣ�ʹת¼���Խ��С�������ʵ�鷢�֣�������pka��ʹ���鵰�����ữ�����ữ�����Ե���������ǿ�����н϶�ĸ���ɣ��������ɵ��鵰���н�ǿ�����������ϣ�ʹ�鵰����dna���룬����鵰��dna�����ֶ�����ת¼��[4,5]
3.7 ���������ϸ����ֳ��ֻ��ĵ���
ϸ������ֳ��ֻ���ϸ��������������������ֳ����ϸ����������dna���ƺ�ϸ�����ѣ���ͨ��ϸ��������ʵ�ֵġ��ֻ���ζ��ϸ���ڵ�dnaͨ��ת¼����mrna���ٽ�һ���ϳ�רһ�Եĵ����ʣ�ø�������յ���ϸ����̬���ṹ��������ɺ��ܵȸ���ˮƽ�ϵIJ��졣ϸ����ֳ��ֻ���һ�Լ�����ϵ����ì�ܵĹ��̣�camp�ڵ������ì����������Ҫ������[13,14] ��camp������ϸ��������ϸ�����ѡ��ٽ��ֻ������á���ˣ�����ʹϸ����camp�������ߵ����ؾ��ܽ���ϸ���������ٶȣ�����ϸ������ֳ�����ٽ�ϸ���ķֻ���[4,5]��������Ϊcamp��ϸ����ֳ����˫��ЧӦ������g0����g1��ʱ��ϸ����ֳ���дٽ����ã�������g1��ʱ�����������á�[15]
���⣬camp��������һ���Ĺ�ϵ����ϸ��������ϸ���е�camp�������в���ģ�������ϸ���ڵ�campһ���������ϸ��ˮƽ��ϸ�����ٻָ�����ϸ��ʱ�����ڵ�campˮƽҲ�漴�ָ�������[4]
4 ��������������ҵ�е�Ӧ��
camp����߶�����������ܺ������ƷƷ�ʡ������û���Ϊ��camp��Ϊ���صġ��ڶ���ʹ�������ø��ʹ��лø������ǿ���Ӷ���ǿ���ڵ��ĺϳɣ����춯��������ٶȣ��յ����أ����������صȣ���ø�ĺϳɣ��ٽ�����ĺϳɴ�л��
4.1 ���������ڼ��������ϵ�Ӧ��
4.1.1 �������������������ϵ�Ӧ��
����������������camp���������õ��������ܺ�����Ʒ�ʵ�Ӱ����һ�¡���ˮƽ��camp������֬����֯��hsl���ʹ֬���ֽ�������ǿ��[16]ͬʱcamp���ɼ����ȵ�����֬��ϸ���Ľ��[17]���ԶԿ��ȵ��ض���֬�����Ĵٽ����á�������ȣ�1992������Ƥ��ע��camp��pde�����Ƽ������������������������������ɣ�����������ʡ��ۼ�����ͺ��˱ȣ����һ�ʹѪ����֬�ʺ�����֬��ø�Ļ������ߡ���ʾ��֬���ͺ�Ѫ֬���߿�����camp������ʹ����֬��ø�������ߡ�������֬��Ա�йأ�����֬�Ĵ����ֽ����������ڽ��͵����ʵķֽ⣬�Ӷ���ǿ�����������ڵij���[18]��
camp���������Ҳ�����ԵĴ��������á���ʿ����1997�����������������������ע��camp�Ʊ�Ʒ�ͱ�Ʒ�Ƽ������������������á�[19]�����µȣ�1993��ע��camp���������������Ե�����Ч����������ʹ��ʱ����ӳ�����Ч�������½���[20]
���⣬camp�����������IJ�ë����ë��ά���ȣ��ı���ë�и��ְ�����ĺ���������ë�����Ļ���������ͨ��������ë�������ڻ�����ë�ҵ�Ӫ����Ӧ��ʵ�ֵġ�[21]
4.1.2 �������������������ϵ�Ӧ��
����ʮ����������ͨ���Դ���С�������ú�������ţ�����ȶ������ٺ�����camp���о�������camp����������������������Ҫ���á����﷼�ȣ�1993�������˲�ͬ�����ɽ�����л�������ı仯������ǰ��������������ߣ�����ά���ڣ������15�죬���е�campŨ��һֱά���ڽϸ�ˮƽ��������߷��ڣ�camp�������ߣ���������ڣ�ֹͣ����ǰ1�������ң���ֻ��������������½������е�camp������½�������Ϊcamp��Ҫ�ٽ���֭���ڣ�cgmp��Ҫ�Ǵٽ���֭���ɣ����߳�һ�������仯��[22]�����ܵȣ�1995����ͬƷ��ţ��Ѫ��campˮƽ����Ϊ������ͬ�ı仯���ɡ�����ǰ��һ�ܳ�С�IJ�����Ȼ��Ѹ���������ڷ����45��ﵽ��ֵ�������90��Ѫ��camp���½������ơ� [23]³��̫�ȣ�1994�����������е�camp�ı仯�������еı仯�෴[24]��shiffield ����ע����Ҷ��أ�camp�ٽ������ķ����ɹ���ʹ��ֲ���������ڵ�ţ������֯�����ͷ���[25]�����о���camp�������������������ɽ������������������ȣ�2002����������һ����Ũ�ȷ�Χ�ڣ�camp������������ɽ��������Ƥϸ����ֳ�дٽ����ã����ʼ����������ԣ���campŨ�ȹ���ʱ�����������á�camp�Ը�ϸ���������ߵ�Ӱ�����ڶ��������ڡ���Դ��camp�ܴٽ�����������ɽ��������Ƥϸ��������֭��ɽ���ҵ��ı�� rillema���������ص�����������camp��ˮƽ��һ��������ԡ�[26]Ȼ��camp����ε�������ϸ�������顢�����ģ�Ŀǰ�в�ʮ��������������������camp�ٽ�����ǰҶgh��prl(����̼���)�����鼤�ص����ɺͷ��ڣ��ӿ���弸��������֯ϸ���Ĵ�л�����ȴ���ƶ��ֶ�����������ϸ��dna��rna�����ǡ��鵰����֬�ĺϳ�[27,28]������������Ϊ��camp�ɽ��ʹ���С�������õ����ٺ��Ǻ��գ�rna�������ǡ���֬���ҵ��ȵĺϳɣ�����һЩø�Ļ��ԣ��������������ɡ�[29,30]������Ϊcamp���������������أ�camp�ٽ���֬�ϳɣ������鵰�ϳɡ�[31]
4.2 ���������ڼ��������е�Ӧ��
��Դ��camp�������ٽ���ά�����м�����������ؼ��ʡ��ȼ��ʡ�����֬�ʣ�������Щ��Ӧ���ż��������Ӷ�����[32]�������ỹ���⼦��л��Ӱ�죬���ɽ������е�֬���������ٽ������ʻ��ۣ��Ը�����֯Ҳ�����Ƶ�Ч��������ʹѪ���ܵ�����֬��������[33]��camp������֬����л�����úͲ����ද��������ͬ��camp�ܴٽ������ද��ļ�������֬ø��hsl���������ߣ��ٽ�����֬���ķֽ��Ѫ֬�ĺ������ߣ��Ӷ��������е�֬����������������Ʒ�ʡ����ڼ����У������������ض����ȸ�Ѫ���أ�������Ҫ��֬�⼤�ء�ţ����ȣ�1998��������Դ�Ի���������⼦���ڵ�gh�ķ��ڡ���л��һ���Ĵٽ����ã������������������Կ��ڶ����������£�������ȴ���ڶ����飬�������camp���⼦����Ѫ��ϵͳ���е��������йأ���һ������Ѫ���е����ʺ����������ڶ����飬�������������й�[34]�����⣬camp����������ߵ����IJ������ܣ����Ƶ��ǵ�Ʒ�ʣ��ɴ��������ľ���Ч��[35]��
5 �йش��ڵ�������Ӧ��ǰ��չ��
Ŀǰ�й�camp����������ϵͳ�о��Ƚ��٣�����Ŀǰ�й�camp���о����������ע��ķ����������ɸ����ģ���ƹ�Ӧ�ô��������ѡ����⣬������Դ��campȷ�����û���������Դ��camp�к�Ӱ��������д��ڽ�һ���о���
��Ȼ���й�camp���о���������ҵ�ϵ�Ӧ�ô����������⣬����Ϊ���������ĵ��ڼ���camp������Դ�㣬��ȡ���Ʊ�������ŵ㡣���⣬�����������������ǵ��������Ʒ�в������������������������쳣��Ŀǰ�ѱ����á���ѡ�ü��صĵڶ���ʹ�����Ӽ��ص�ЧӦ���ȿ�����߶����������ܺ������ƷƷ�ʣ������κβ���ЧӦ����ˣ������camp���о����������������е�Ӧ�þ������õ�ǰ����
�ο����ף�references��
[1] sutherland e m, ye i & butcher r w. the action of epinephrine and the role of adenyl cyclase in hormone action [j]. recent progress in hormone research, 1965,21,623~646
[2] ������. ϸ����Ϣ�����[m]. ����������ҽ�ƴ�ѧ���й�Э��ҽ�ƴ�ѧ���ϳ����磬1998��168~176
[3] ����ɭ����Ԭ־��.�����������������ڴ�л���ɵ��о�[j]. ������ҽѧ����2000��31��2����124~130
[4] ����. camp��ϸ����л�еĵ�������[j].���ﻯѧ������������չ��1981��3��11~14
[5] ���ҵ�������֣�������. ϸ���ź�ת��[m] (�ڶ���). ��������ѧ���������磬1997��52~70
[6] gerosa ma. cyclic nucleotide deesin meduloblastomas correlative study of tumoral and cerebrospinal fluid levers [j]. neurosurgery, 1980, 7:359
[7] dana s, et al. induction of human growth hormone promoter activity by the adenosine 3��,5��-monophsphate pathway involves a noval responsive element [j]. ca., 1989,17(3): 72~75
[8] ganced j m, adelstein r s, morgan k g et al. regulations by cyclic nucleotides and phorbol ester of calcium efflux form cultured bovine adrenal chramaffincells [j]. tibs, 1985,10:5
[9] ���ͣ�ţ����ʯ���۵�. �����������⼦�ڷ��ڼ����߹��ܵĹ�ϵ[j]. �й���ҽѧ����2001��21��3����278~280
[10] monall e. expression of myosin and action in e. coli. merhods, enzymot, 1991, 196:368~389
[11] montminy m r, sevatino k a, wagner j a, et al. identification of a cyclic amp-responsive element within in rat somatostation gene [j]. proc natl acad sci usa, 1986,83:6682~6686
[12] montminy m r, bilezkjian l m. binding of a nuclear protein to the cyclic-amp element of the somatostation gene [j]. nature, 1987,328:175~178
[13] burgering bmt, bos jl. regulation of ras-mediated signing: more than one way to skin a cat[j]. trends biochem sci, 1995a,20:18~22
[14] dumont je, jauniaux jc, roger pp. the cyclic amp-mediated stimulation of cell proliferation [j]. trends biochem sci, 1989,14:67~71
[15] miyazaki m, wahid s, bai l et al. effects of intracellular cyclic amp and cyclic gmp levels on dna synthesis of young-adult rat hepatocytes in primary culture [j]. exp cell res, 1992, 200: 404~409
[16] ecan j j, greenberg a s, chang m k et al. mechanism of hormone-stimulated lipolysis in adipocyles: translocation of hormone-sensitive lipase to the lipid storage droplet[j]. proc. natl. acad. sci. usa,1992,89:8537~8541
[17] liu c y and mills s e. decreased insulin binding to porcine adipocytes in vitro by beta-adrenergic agonists[j]. j sci, 1990,68:1603~1609
[18] �����壬�ν�����³��̫����Դ����Դ��camp����������֬�ʺ͵����ʳ�����Ӱ�죮����ũҵѧ����1992��1(4)��19?22
[19] ��ʿ��. ��״����������������Ч�����о�[j]. ����������ҽ��1997��6��14~16
[20] �����£���ʿ������֮����. ��״�������������������Ч����̽[j]. �ຣ������ҽ��־��1993��23��3����23~24
[21] black j l, reis p j, ed. physiological and environmental limitation to wool growth[j]. armidale; university of new england publishing unit,1979,337~352
[22] ���﷼��ŷ�����죬��־��. ��ͬ�����ɽ�����л�������ı仯[j]. ����ũҵѧ����1993��2��4����81~84
[23] �����ܣ��ž��䣬�����ٵ�. ��ţѪ����������ˮƽ��������ܵĹ�ϵ[j]. ����ũҵ��ѧѧ����1995��17��4����60~62
[24] ³��̫���ξ��ޣ�������. ��������������[j]. �й���ţ��1994��6��38~40
[25] sheffield lg.. influence of cholera toxin (an adenylafe cyclase activator ) on deoxyribonucleic synthesis of bovine mammary tissue in vitro and in athymic nude mice[j]. j dairy sci. 1989,72(1):41~48
[26] rillema j a. cyclic nucleotides and the effect of prolactin on uridine incorporation into rna in mammary gland explants of mice[j]. horm. metab res, 1975,7:45
[27] clayto r n. cyclic adenosine nucleotides and growth hormone-releasing factor increase cytosolic growth hormone messenger rna levers in cultural rat pituitary cells[j]. j edocr, 1986,110:51~53
[28] mature r a. transcriptional regulation of prolation gene by ergocription and cyclic amp[j]. nature, 1981,294:94~97
[29] perry j w, oka t. cyclic amp as a negative regulator of hormonally induced lacto genesis in mouse mammary gland organ culture[j]. prot natl. acad. sci usa,1980,77:2093~2097
[30] sheffield l g. cyclic nucleotides concentrations and protein kinase activities of bovine mammary tissues maintained in athymic nude mice: effects of mammogenic and lactogenic hormone[j]. j. dairy sci, 1988,71:75~83
[31] �ԺƱ�³��̫. �������������ָ����������ܹ�ϵ���о�[j]. ����ũҵ��ѧ��1992��2��21~22
[32] �Ÿ�÷��Ѧ���������. ��Դ�Ի���������⼦�������ܵ�Ӱ��[j].�й�������־��2001��37��3����25~26
[33] �Ÿ�÷�������ƣ����µ�. ����������⼦ѪҺ����֯����ָ���Ӱ��[j]. �й����ݣ�2000��22��12����10~11
[34] ţ���ᣬ�������������ĵ�. ��Դ�Ի��������aa�⼦����������Ӱ��[j].�й���ҽѧ����1998��18��3����299~301
[35] ţ���ᣬ�����ѣ��³�����. ��Դ��camp�Ժ�������Ʒ�����������ܵ�Ӱ��[j].��������ҽ��2000��32��5����17~18
camp and its application in animal science
wang shanfeng��chen anguo
��animal science college, zhejiang university��
abstract as an important intracellular second messenger in cells, cyclic adenosine monophosphate is involved in cellular differentiation, cellular proliferation, cell metabolism and immunoreactions. moreover��it plays an important role in the above process. in the article, physiological function of camp and its application in livestock husbandry are reviewed. finally, the correlative problem and the prospect of application of camp in animal science are mentioned.
keywords: camp, physiological function, animal science.
����飺
���Ʒ棬�У���1976?�����㽭��ѧ�����ѧѧԺ2001��˶ʿ�о���
�°��������ڣ�˶ʿ�о�����ʦ��ͨѶ��ַ�뵥λͬ��һ����
TAG:
����